Balfourier F, Bouchet S, Robert S, et al. Worldwide phylogeography and history of wheat genetic diversity[J]. Science advances, 2019, 5(5): eaav0536.
通过采用high-density Affymetrix Axiom SNP array对4506个landrace和cultivar进行基因分型,作者获得了一个包含280226个SNP的数据集,并在进一步筛选后得到了113457个高质量SNP。这些高质量SNP包含99,333个多态性高分辨率(PHR)双等位基因SNP和14,124个脱靶变异体(OTV)(同时检测核苷酸多态性和存在缺失变异的标记)。通过IWGSC v1.0,作者确定了这些SNP在基因组上的位置。
单倍型分析
单倍型分析将基因型解析为8741个较大的区域。作者用单倍型块进行样本距离计算发现,相对于直接用SNP,单倍型算出的相似度低很多。作者认为是等位基因数量更多(尤其是稀有的)导致单倍型算出的相似度较低,并猜想这或将有利于展示SNP难以追踪的近期的变异。单倍型块的数量与每个染色体中SNP总数高度相关。每个单倍型块的类型总数均值为4,在2-20之间。单倍型块的中位大小为105
Kb,并由85%的块小于1Mb,均值863 ± 4595 kb。这个巨大的SD反映了小麦染色体的结构划分。
单倍型计算中,首先用Beagle软件估计缺失的数据,并删除丢失数据超过5%的行。使用PLINK中的Haploview算法区分单倍型。In each haplotypic block, 95% of pairwise measures of marker LD (D′) were superior to 0.98.
群体结构
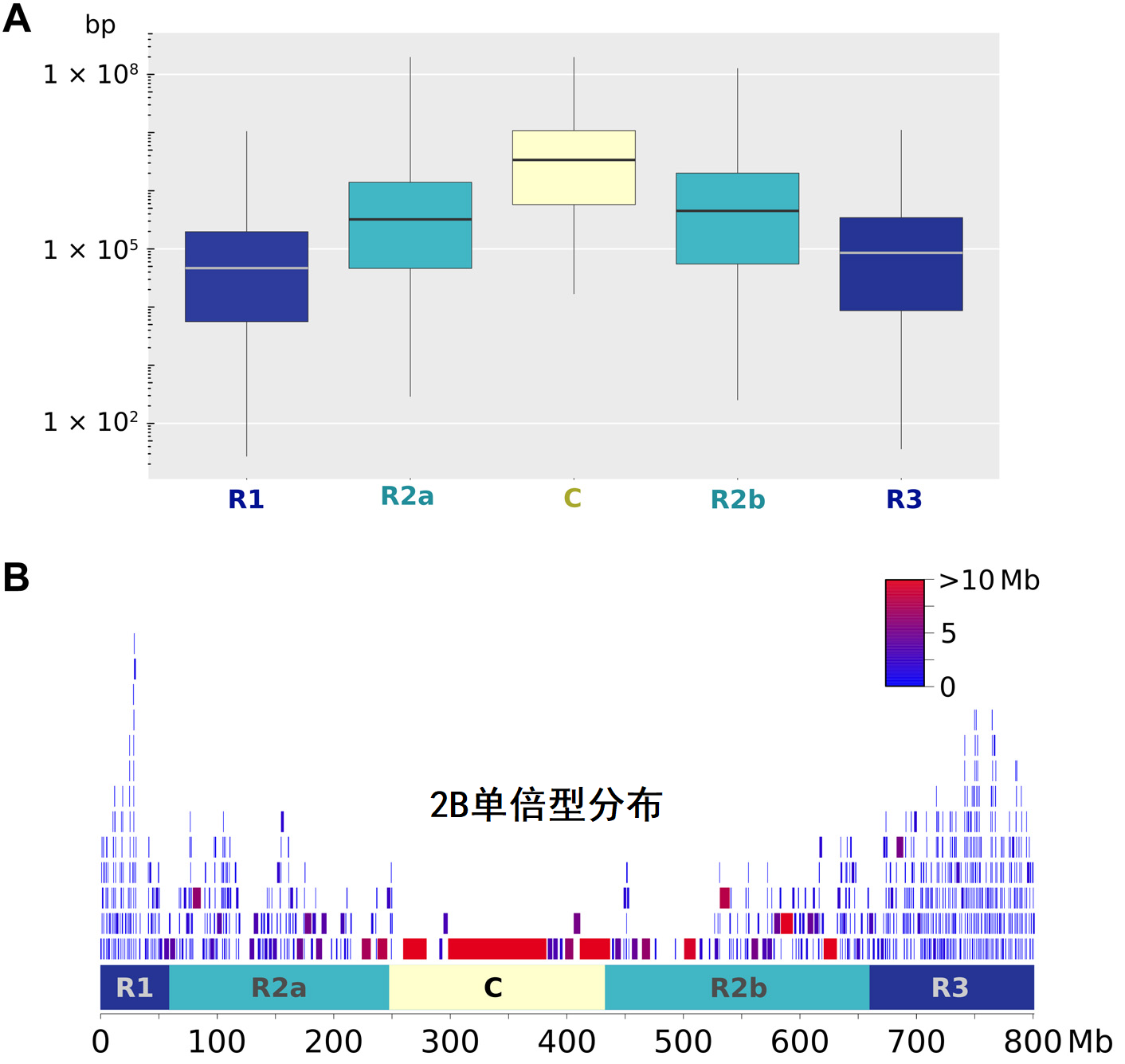
STRUCTURE分析显示,K=6至9时分组更为稳定。作者接着介绍了K=2至K=8时的分组概况。下图中,对角线显示了分组内稀有等位基因数量(频率<0.05),对角线上方是(F_{ST}),下方是两个种群重叠的稀有等位基因数量。
(F_{ST})采用hierfstat R package计算。
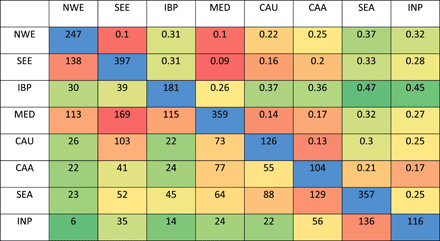
地理映射
PCoA的第一轴和第二轴地理投影显示,存在两个分化极,一个在欧洲,一个在亚洲。它们都在地中海地区汇聚,这与有关将小麦驯化置于新月沃地的文献一致。距新月的遗传和地理距离之间的关系支持这样一个事实,即有两个分化源于一个共同的中心:新月(图S3)。通过对稀有等位基因的研究,作者认为,小麦的扩张可能始于地中海地区。然后,面包小麦在新的环境中沿两条主要的人类迁徙路线进行了分化,从新月沃地走向欧洲(沿多瑙河路线向北(SEE)和向西(NWE))以及沿丝绸之路走向亚洲。
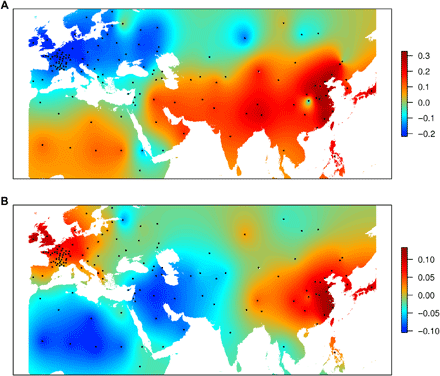
PCoA采用Kriging method得出。The spatial prediction is based on a Gaussian model.软件采用了TESS3。
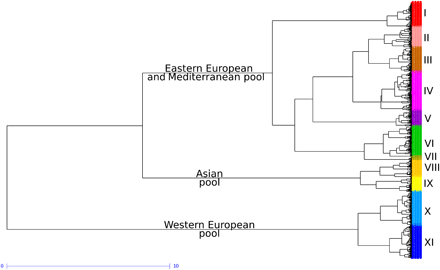
系统发育分析与单倍型分析
系统发育树显示了三个大基因池。东欧和地中海的基因池中有包含了94%的SEE, MED, and IBP landraces,其中包含所有传统和现代多样性中83%的、不是来自北欧和西欧的部分。现代品系和亚洲地方品种之间的高分化指数(SEA和INP;平均(F_{ST}) = 0.35)支持以下事实:原始的亚洲多样性库对现代品种没有太大贡献。同时,作者发现北欧和西欧的育种计划主要使用当地的地方品种。
通过各国、各地区主要单倍型群(欧洲、MED、中东与中亚、东南亚、中国、非洲等),作者讲述了各地育种的故事。

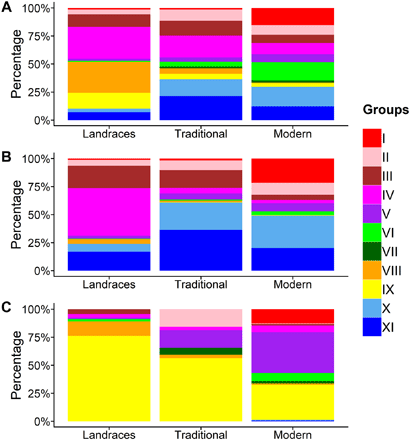
下图展示了各品种(时期)间遗传多样性的改变。图中,A是世界范围,B是欧洲范围,C是中国范围。
结构变异与小麦育种
对OTV分布的详细分析显示,大于5 Mb的80多个SV块,累计长度大于4600 Mb,即几乎占整个面包小麦基因组的三分之一(表S5)。这些大块可能对应于外来的基因渗入,如1BL/1RS易位、[偏凸山羊草2NS易位染色体上2A](https://doi.org/10.2135/cropsci2003.1839)等。但由于本文方法原因,小外来片段(称为cryptic introgressions)难以被检测。因此,当前小麦种质中存在的外来DNA总量可能远远大于该数量。
由于驯化和选择,改良材料中的SNP自然遗传多样性显着低于在地方品种中观察到的SNP自然遗传多样性。然而,与本地品种相比,现代品种中丢失了9%的等位基因,而现代品系中发现了8%的新等位基因。这些现代基因特有的等位基因中有19%含有OTV,而地方品种特有的等位基因只有7%。这表明育种计划带来了一些与外来基因渗入有关的富含SV的新等位基因,从而导致总体遗传多样性与地方品种和现代品种相似。
作者对面包小麦的介绍
面包小麦(Triticum aestivum L.)是一种异源六倍体物种,起源于连续两轮杂交。第二个杂交事件被认为是新石器时代期间都发生在新月沃地,〜8000至10000年前(1,2)。然后,面包小麦种质沿着古老的人类迁徙道路进化。从现在开始,该地区的第一批农民从8500到2300一直向西传播到欧洲,再向东传播到亚洲(3)。在欧洲和亚洲传播后,驯化的小麦种群已适应当地环境,成为所谓的地方品种。从16世纪开始,面包小麦被引入新世界,首先是在拉丁美洲,然后在北美和澳大利亚(3)。在过去的两个世纪中,在欧洲和亚洲组织了育种计划,以改善这些地方品种。最后,第二次世界大战后,绿色革命期间农作物(特别是小麦)中矮化基因的引入,导致了全世界基因库的显着改变(4)。如今,小麦每年生产超过2.2亿公顷,近750公吨,是世界上耕种和消费最多的农作物之一,每天提供15%的卡路里消耗。自从狩猎聚会向农业过渡以来,面包小麦对文明的兴起至关重要。通过反复选择来适应人们的需求并适应不同的环境。在这里,我们报告了一项全球植物地理学研究,旨在了解小麦传播和分化的这一复杂历史。
样本
4506个品系选自Institut National de la Recherche Agronomique (INRA) Biological Resources Center收集的12000个品系。选择结合了前人的研究。
每个accession的详细信息见Data file S1
基因分型
作者通过Affymetrix GeneTitan system平台执行了分型操作,并通过修改过的Affymetrix Power Tools 和SNPolisher来执行allele calling环节,以适配小麦基因组的特点。对于所有SNP,作者计算了HomRO和HomFLD,并以3.6作为HomFLD阈值进行过滤。数据处理上,探针杂交之后,没有通过质量检查的SNP被重新处理。被SNPolisher分入OTVs的探针集被OTV_caller进行了两步分析。SNP被Affymetrix software通过cluster patterns分为6组:PHR, OTV, monomorphic high resolution, no minor homozygous, call rate below threshold, and others。只有 PHRs and OTVs被用于后续研究