一、一些基础概念及方法
[1] Vellend M. Conceptual synthesis in community ecology. Q Rev Biol. 2010 Jun;85(2):183-206. doi: 10.1086/652373. PMID: 20565040.
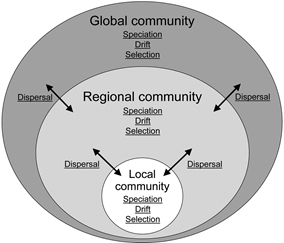
群落生态学的概念相对mess,这篇文章梳理了成种、漂变、选择和扩散在群落动态中的作用。
[2] Cavender-Bares J, Kozak KH, Fine PV, Kembel SW. The merging of community ecology and phylogenetic biology. Ecol Lett. 2009;12(7):693-715. doi:10.1111/j.1461-0248.2009.01314.x
谱系相关的经典综述,被引 1142
[3] Wilcox TM, Schwartz MK, Lowe WH. Evolutionary Community Ecology: Time to Think Outside the (Taxonomic) Box. Trends Ecol Evol. 2018;33(4):240-250. doi:10.1016/j.tree.2018.01.014
竞争与进化。达尔文的相似性竞争假说有两个比较大的问题,一是物种共存不仅取决于生态位相似性也取决于竞争能力相似性,二是谱系关系不能很好预测一些性状(性状进化速率非恒定,且存在趋同进化)。
[4] Violle C, Enquist BJ, McGill BJ, et al. The return of the variance: intraspecific variability in community ecology. Trends Ecol Evol. 2012;27(4):244-252. doi:10.1016/j.tree.2011.11.014
种内变异。模仿群体遗传学里的F统计,提出了基于性状的T检验(T指Trait)。未来方向(目前的困难):(1)多尺度的采样策略;(2)考虑时间变异;(3)基于genetics的方法和基于traits的方法的统一。
[5] Damgaard C. A Critique of the Space-for-Time Substitution Practice in Community Ecology. Trends Ecol Evol. 2019;34(5):416-421. doi:10.1016/j.tree.2019.01.013
空间代替时间方法的局限性,尤其是在非稳定的环境中(比如黄河三角洲)。空间变异,只能分析和某个生态过程相关,或者获取时间序列数据。
[6] Mouquet N, Devictor V, Meynard CN, et al. Ecophylogenetics: advances and perspectives. Biol Rev Camb Philos Soc. 2012;87(4):769-785. doi:10.1111/j.1469-185X.2012.00224.x
(谱系关系-生物互作网络。Beyond the fundamental question of how evolutionary history contributes to shape communities, ecophylogenetics will help ecology to become a better integrative and predictive science.)
二、热点之多营养联动
[1] Seibold S, Cadotte MW, MacIvor JS, Thorn S, Müller J. The Necessity of Multitrophic Approaches in Community Ecology. Trends Ecol Evol. 2018;33(10):754-764. doi:10.1016/j.tree.2018.07.001
相比以往的单营养级或单类群研究,多营养级研究得出的结论会相同吗?在实际调查中,是优先考虑营养级的全面,还是分类类群的全面?里边提出的科学问题可以借鉴。
[2] Warne RW, Baer SG, Boyles JG. Community Physiological Ecology. Trends Ecol Evol. 2019;34(6):510-518. doi:10.1016/j.tree.2019.02.002
动物内稳态控制(homeostatic control)对生态系统的级联影响。微生物角色的处理。突破关键在于,研究方法整合药物投放(改变动物内稳态)、高分辨率追踪技术、稳定同位素技术。
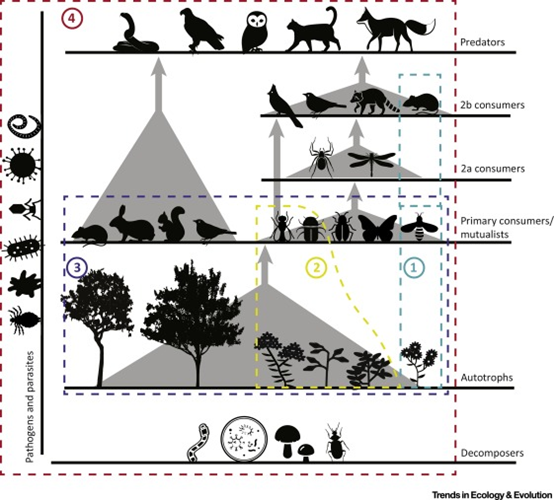
[3] Laforest-Lapointe I, Paquette A, Messier C, Kembel SW. Leaf bacterial diversity mediates plant diversity and ecosystem function relationships. Nature. 2017;546(7656):145-147. doi:10.1038/nature22399
揭示现象,某营养级的功能也受其他营养级多样性的影响
三、热点之微生物群落
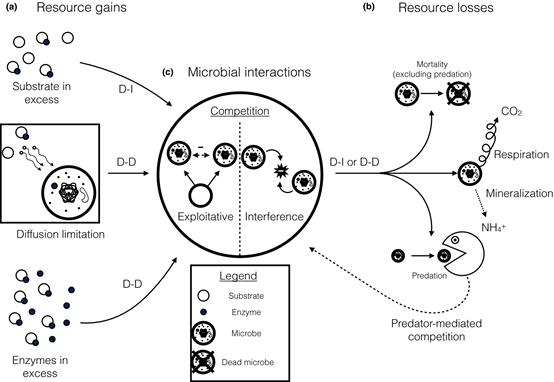
[1] Buchkowski RW, Bradford MA, Grandy AS, Schmitz OJ, Wieder WR. Applying population and community ecology theory to advance understanding of belowground biogeochemistry. Ecol Lett. 2017;20(2):231-245. doi:10.1111/ele.12712
微生物互作对生态系统的功能。相比环境-生物互作对生态系统功能的研究,群落动态和生物互作对生态系统功能的研究较少。
[2] Gibbons SM. Microbial community ecology: Function over phylogeny. Nat Ecol Evol. 2017;1(1):32. Published 2017 Jan 4. doi:10.1038/s41559-016-0032
后一篇是前一篇的编者按,写得非常精彩。以宏观生态下,各个大陆由于进化和隔离的原因,物种差异很大,但功能相似。Louca发现了凤梨微宇宙体系里微生物的功能可以保持稳定,但存在物种周转(分类多样性差异很大)。这和功能性状不是谱系保守的分析结果是一致的。物种分类差异大,可能不是扩散限制的原因(不符合地理距离隔离模型),可能是植物生理和环境差异造成的(虽然已经测量的环境因素只能解释5%)。研究的不足之处,是测得是16S,功能是预测的,没有测更多直接生态功能相关的指标。后续科学问题:生物群落物种组成不同,但生态功能相同的原因。
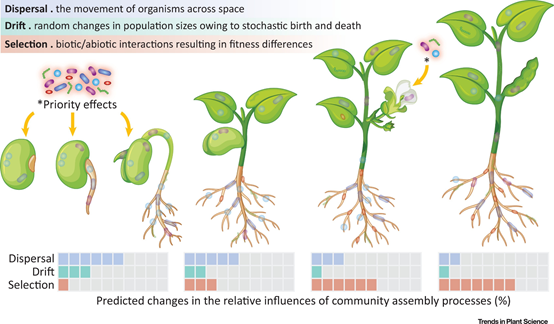
[3] Dini-Andreote F, Raaijmakers JM. Embracing Community Ecology in Plant Microbiome Research. Trends Plant Sci. 2018;23(6):467-469. doi:10.1016/j.tplants.2018.03.013
群落构建由选择、扩散、漂变、成种等因素影响。微生物以往以选择作用研究为主,需要把群落生态学框架转移到微生物组的研究中。
[4] Prosser JI, Martiny JBH. Conceptual challenges in microbial community ecology. Philos Trans R Soc Lond B Biol Sci. 2020;375(1798):20190241. doi:10.1098/rstb.2019.0241
写作值得学习。概念上有四个需要考虑的问题,1、进化;2、微生物互作;3、空间(尺度);4、响应环境变化及生态功能。落脚点是生态系统功能。
四、群落构建与生态功能的耦合机制
[1] Aponte, C, Kasel, S, Nitschke, CR, et al. Structural diversity underpins carbon storage in Australian temperate forests. Global Ecol Biogeogr. 2020; 29: 789– 802. https://doi.org/10.1111/geb.13038
以上两篇文献主题一致。互作效应、选择效应;结构多样性,基于树型大小的变异,可以区别于功能多样性,也受林龄的影响
[3] Bannar-Martin KH, Kremer CT, Ernest SKM, et al. Integrating community assembly and biodiversity to better understand ecosystem function: the Community Assembly and the Functioning of Ecosystems (CAFE) approach. Ecol Lett. 2018;21(2):167-180. doi:10.1111/ele.12895
丰富度-组成两步模型,丢失-获取-改变三部模型(群落构建模型),五步模型(整合前两个模型)
[4] Cadotte MW. Functional traits explain ecosystem function through opposing mechanisms. Ecol Lett. 2017;20(8):989-996. doi:10.1111/ele.12796
选择效应和互补效应的大小取决于性状相似性;生态位(niche)决定物种共存,适合度(fitness)决定物种排斥,二者测度性状?多个,单个?突出功能性状在物种共存/排斥、生态功能中的桥梁作用。展望:多系统,种内变异;意义:功能最大化的设计。
[5] van der Plas F. Biodiversity and ecosystem functioning in naturally assembled communities. Biol Rev Camb Philos Soc. 2019;94(4):1220-1245. doi:10.1111/brv.12499
非实验系统(自然系统存在群落构建过程)中的BEF研究的最全面综述。
[6] Godoy O, Gómez-Aparicio L, Matías L, Pérez-Ramos IM, Allan E. An excess of niche differences maximizes ecosystem functioning. Nat Commun. 2020;11(1):4180
通常会物种共存理论中的生态位分化来解释生态功能的互补效应和选择效应,但未有实证。本研究在生产力功能上,验证了物种共存和生态功能的驱动力是相同的。
五、其他