为进一步了解这种连接性产生的差异,在认识soma这个独立的逻辑单元后,进化过程设计了一整套系统设计来使用这个逻辑单元。为促成细胞体之间发生连接构成系统dendrite和axon之间的连接需要引入新的功能单元Synapses。Synapse叫做突触,书上说得特别好,这玩意是特指细胞间通讯的部位,它本身不是一种细胞,而是细胞之间的连接点。
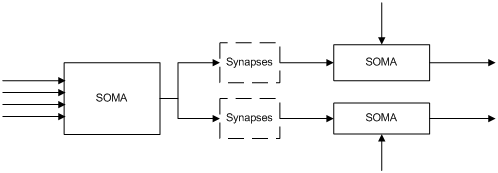

输入和输出的信号沿着神经纤维一直传递,这种单一材质内的活动(这种信号估计是离子迁移,我看的资料没咋说清楚)按照最快120m/s的速度在Dendrite和Axon中传递。 只观察神经微电路的场景下Dendrite和Axon就被统一,简化为连接(对我这种怕麻烦的人就视其为导线),整个连接结构就被简化为仅有cell Body和synapses的简单构成,这样我们就获得了细胞之间形成复杂网络的基础要素。这就算齐活了!下面是不是可以往上堆功能了?不急,我们再耐心听完synapase的故事。Synapses是在cell body之间的连接节 点,它本身不是简单的连接器,而是具备功能的处理单元。 对于普遍的情况信号的传递跑到synpase(接头处)就停了下来,信号通过synpase中的化学介质传递到下一个细胞体。这种叫做化学传递,这种信号传递方式细节有点小复杂,我这里就跳着简单说一下。
端上有一系列Ca+离子,当soma被激活时,产生的点位差会导致Ca+离子的活动。当Ca+离子浓度增加会导致vesicle跑到synpase的前端破壁而出(原来的说法更形象:vesicle向间隙"吐"递质),向细胞间隙中释放一些称作递质的化学物质(谷氨酸/多巴胺这一类的玩意,种类大于100我记不住),整体上就是这些递质会从synapses内跑到细胞间的接缝处形成聚集(对应下面右图中的小点)。接收端在对应的位置有一堆ions通道,在递质发生一通化学反应后,这些离子通道会被打开,(可认为递质的一个重要作用是打开离子通道的钥匙)离子通过接收信号的细胞的离子通道接收离子流。这些离子从而影响细胞的突触后电位(EPSP)。而一个细胞体所连接的输入是来自于多个输出的。用物理常识就能知道细胞体的电压特性是这些输入的总和结果。因此这些突触整体上表现出相加的关系。当累计电位小于阈值时(就是动作电位),细胞就不激活,反之,如果超过动作电位,而且本身不处于充电状态(Repolarization),这些输入就会触发axon发出一个动作脉冲。
貌似这段粗糙的解释就说明了synpase从电->化学->电的传递过程,深入进去这其中还包含一个递质的量子释放过程,和递质的回收、分解的过程。出于降低脑容量需求的目的,个人直接忽视这两个重要的环节。好在这部分不影响到后面要构建的基本内容,这就直接跳过拉倒。
这种诡异的方式使得信息传递只能单向进行,真真切切的起到了隔离的效果。同样的,由于传递本身是由于化学离子的移动进行,这些离子在传递完信息后会被回收。Ca+就来回跑圈,刺激递质的释放,就达到了传递信号的目的。同时也看到这里面存在两个延迟因素,一是信号传递的延迟和电压缓慢上升,第二是回收周期中延迟,电压的缓慢下降。按照书上的说法这种传递会产生1ms左右的延迟。另外一方面的影响是,Ca/Na/K离子的浓度会影响形成的电流(说白了就是离子通道内通过的正离子总数),离子浓度由synpase上的vesicle内的递质总量决定。对于一些特定的情况,前后的细胞间的synpase是存在物理的Gap Junction的。离子就直接跑过去这种情况就显得简单很多。对接收端而言,接收端提供了一条离子通道和间隙连接,而对于抑制脉冲(负脉冲的情况),CI离子会流入(就是盐巴里面的氯),这玩意是负离子从而拉低接收端的电位,整体上要求其他synpase需要输入更多的正离子,起到事实上拉高动作电位的作用,从而体现为抑制。
看到这里感觉补钙和吃盐都很重要啊。
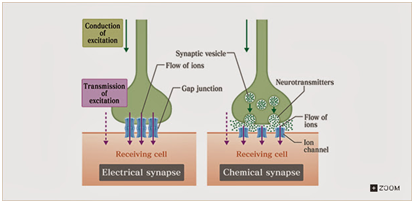

从上面的这堆罗里吧嗦的描述可以总结synpase的几个特性。
首先,synapses具备类似二极管的特性,使得信号传递单向的进行,即是下一级单元的点位状态不要影响上一个信息输入单元。这让我们能在图上画出来具体的箭头方向。
第二: 对化学类synpase,会产生延迟。
第三: 对细胞体的电位而言,这些synpase的输入不是单个影响细胞整体,而是一系列输入的累加的总体表现。因此输入synpase之间总体上表现为加法输入(对负电位的脉冲输入则体现为加个负数嘛)。
第四:动作电位的影响能力决定于发送端的递质浓度和接收端的离子通道数量,这貌似是一个可以逻辑上调节连接拓扑的手段。
自于vesicle里面的递质从哪儿来的,这里简单说两嘴:
1.大分子的递质是细胞体自己合成的,合成好了以后会装到囊泡里面,慢慢(大概400m/天的速度)搬到端上的synpase。当然,运输的代价就是消耗ATP。搬过来以后如果遇到激活情况,一把就用掉。
2.小分子的递质就比较先进,细胞核把RNA从DNA上拷贝出来,把RNA直接带到synpase,在synpase再用RNA组装递质。显然这个模式比较先进,比较节能。
上面说的这些信息由于发现和确认都比较早普遍在90年代就全部考证玩了,在普通的神经学1年级教材都能找到。下面被迫引入点高级货,所谓高级货就是新搞出来的和还没看到人说清楚的,也不知道是不是普遍认可的东西,引入实在情非得已,后面我们需要啊。
核心问题是synpase拓扑连接的问题,如果只是往复信息传递玩激活游戏,无论怎么设计连接结构在拓扑结构上都会与冯式计算机陷入同样的问题、整个信息通路的基本连接关系是在这个基础上无法产生根本性改变,所有的处理通路在发育完成后就固定了能修改的只是信息的输入。
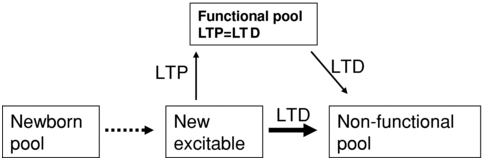
1973年Terje Lømo搞了个对兔头的研究,发现兔头海马体上的synpase存在一个叫做LTP的现象。这LTP具体是什么东西,简单的说就是通过长时间对兔子的特定输入通路上连续一段时间的加载人工产生的电信号(100Hz 脉冲),对细胞体反复激活,发现对应的synpase产生的等效输出竟然提高了(当然输出是无法测量的,他们是测量接收信号的细胞体上的EPSP).这现象说明synpase很可能具备"熟练度"的特性。这开启了对Synaptic可塑性的研究,过了几年日本人Masao Ito又发现了一个叫做LTD的现象.请看下图,基本就是激活时间越长,输出的电压越强烈。这些实验观察证明了一个可以理解为synpase可被特定情况调整的基本逻辑。而这个逻辑与40年代Donald Hebb 搞出来一套叫做Hebbian learning的假说-多用就变强基本符合。
--------------------------------------------------------------------------------------------------------------------------------


狂读一通后对这个LTP摸了基本情况,简单说LTP现象是这样的。对一个突触而言,突触可分为释放化学递质的突触前,和接收化学递质的突触后。突触后LTP在2000年左右似乎搞得比较清楚了,突触后接收递质的受体有两类NMDA和AMPA,NMDA是固定的离子通道,当多次信号输入后,其周边的正离子浓度会提高。在较高的离子浓度下会促成AMDA的产生,相当于在突出后添加了新的离子通道。在同等递质浓度下,离子穿透进入的可能性就得到提高,从而形成“熟练”减强的表现。由于一个通路(相同的两个soma间)上由具体多个synpase连接都发生类似的现象,因此可以观察到接收能力渐强的现象。


LTP对于突触前也确定观察到类似的反复发送后增强的机制,只是似乎目前没有完全搞清楚原理。2011年左右Nisticò et al倒是提了套机制,我本身不是他们这个圈的,不知道普遍认不认这套说法,我就不乱说了。贴个Nistico的图出来,有兴趣的可以瞄一眼。

LTD是个逆向的过程,整体面向抑制过程。与LTP类似,LTD好些材料都要有nature的会员才能看,没渠道也就了解个框架。按照J Physiol. 的说法LTP和LTD本质上就是激活离子通道和反激活离子通道的过程。

------------------------------------------------------------------------------------------------------------------------------
把横线中间的内容简单一归纳,就可以详细说下第四条特性的具体表现:
LTP效应:
激活细胞体,如果接受细胞也被激活的话,synpase的接收增益就会提高。
激活细胞体,synpase的发送增益也会提高
LTD
反之如果出现超极化,目标细胞在被抑制的情况下被激活的话,对应的synpase接收增益会降低。
LTD对发送增益的问题没搞清楚,不乱整。
简单总结为,synpase 会随着soma激活和接收soma的激活情况的而改变其电位信号的增益(其实是衰减比例了)。
由此我们总算在整个系统组织单元内找到一个可以将信息持久化的单元,这玩意对整体功能产生质的影响,简单说就是终于看到信息存储单元了。就像DDR和FLASH一样的存在。具体怎么搞,我们后面细说。
至此,在已经了解基本单元特性的前提下.可以按照理解的特性开始对机器学习算法进行粗暴的观察,以试图在两者之间相互印证。这两个小节的知识对Deep Learning之前的学习算法而言基本已经足够支撑。对于HTM和Deep Learning的会用到皮质整体生物特性和针对pyramidal的细胞特点,在后面专门分开两个章节的笔记进行梳理。以避免在不必要的阶段引入太过复杂的内容。